Phytochemicals, Nutritional Value, and Antimicrobial Properties of Xanthosoma mafaffa
-
Kanabiya Essodjolon Prospère

Department of Chemistry, Faculty of Sciences, University of Lomé, Lomé, Togo
Novidzro Kosi MawuénaDepartment of Chemistry, Faculty of Sciences, University of Lomé, Lomé, Togo
Dotse KokouviDepartment of Chemistry, Faculty of Sciences, University of Lomé, Lomé, Togo
Koumaglo Kossi HonoréDepartment of Chemistry, Faculty of Sciences, University of Lomé, Lomé, Togo
Canabady-Rochelle LaetitiaUniversity of Lorraine, French National Centre for Scientific Research, Reactions and Process Engineering Laboratory, F-54000 Nancy, France
| Received 16 Feb, 2025 |
Accepted 15 May, 2025 |
Published 16 May, 2025 |
Background and Objective: Xanthosoma mafaffa is a food plant used in traditional medicine in Togo to treat anemia and several infectious pathologies. The present study aims to valorize this species through the study of its biochemical compounds and their antimicrobial activities. Materials and Methods: The powders of young or mature leaves of X. mafaffa were macerated at a solid/liquid ratio (10% w/v) at ambient temperature for 72 hrs in a hydroethanolic solvent with an ethanol/water ratio of 50/50 v/v. The biochemical compounds of nutritional interest of young or mature leaves of X. mafaffa were determined by the usual AOAC methods. The antimicrobial activity of the extracts was carried out against Gram+ and Gram‾ bacteria using the agar diffusion method. All biochemical and antimicrobial experiments were conducted in triplicate, with results expressed as Mean±SEM. Statistical significance (p<0.05) was analyzed using DMRT in SPSS 11.5. Results: The contents of biochemical compounds were 38.8±0.4 and 30.7±0.7% for digestible carbohydrates; 24.3±1.3 and 30.4±2.2% for fibers; 17.1±1.7 and 15.8±0.6% for proteins; 3.2±0.4 and 3.4±0.6% for lipids for young and mature leaves, respectively. The presence of Na, K, Ca, Mg, P, and Fe in the leaf samples, at levels deemed significant, is confirmed by spectrophotometric analysis, which also revealed Na/K ratios <1 and Ca/Mg and Ca/P ratios >1. Conclusion: The results underscore that the nutritional composition of X. mafaffa leaves is comparable to that of vegetables and legumes frequently included in daily meals. The young leaves extract obtained with the 50% v/v ethanol/water ratio showed an appreciable inhibitory activity against Klebsiella pneumoniae.
INTRODUCTION
The emergence of diseases around the world makes developing countries more vulnerable. Infectious diseases represent a major public health challenge in these countries, due to their prevalence and considerable health impact1. Indeed, they cause more than 17 million deaths per year worldwide, with over half of these occurring on the African continent2. The pathogenic agents at the origin of these infections are varied and include fungi, bacteria, protozoa, and viruses. The discovery of antibiotics marked a significant advance in the medical field, leading to a notable reduction in the incidence of infectious diseases3. However, the uncontrolled use of these molecules has led to resistance phenomena of many infectious agents4. In developing countries, for cultural reasons and economic difficulties, populations have always used plants to treat themselves. Also, nowadays, with the ever-increasing cost of available drugs, associated with the emergence of multi-resistant pathogens, there is a renewed interest in the African pharmacopeia5.
Furthermore, the ongoing issue of malnutrition represents a major public health dilemma, as inadequate vegetable consumption in unbalanced diets is believed to account for roughly 31% of ischemic heart disease and 11% of stroke cases6. This situation is intertwined not only with the quantity and quality of food intake but also with the limited information and exposure to local resources. Given the resistance of microorganisms to anti-infective treatments, it is essential to pursue novel bioactive substances for effective patient management7. Plants have been used since prehistoric times by humans for nutritional and therapeutic purposes. Indeed, some edible vegetables that enter the diet of the population abound in pharmacological properties8. Vegetables are an important sources of alkaloids, flavonoids, tannins, carotenoids, and especially vitamins, and thus possess several properties such as antioxidant, anti-inflammatory, antimicrobial, and anticancer activities9. As a result, their consumption contributes to the improvement of nutritional status and the prevention of pathologies in rural and urban areas. By way of illustration, therapeutic applications include the use of Parkia biglobosa fermented seeds against hypertension, Adansonia digitata leaves and pulp against anemia, and Sclerocarya birrea pulp against nutritional deficiencies, as traditionally practiced10. Similarly, Moringa oleifera leaves are utilized for their significant nutritional attributes and demonstrated therapeutic potential11,12. To exemplify, they demonstrate anti-inflammatory actions that could alleviate arthritic symptomatology, exhibit antioxidant effects that defend against oxidative stress, and may aid in the reduction of blood glucose concentrations, thereby providing therapeutic potential for diabetes management12. All over the world, and especially in many African countries, edible plants are used as food and thus contribute to meeting the nutritional needs of the population13. In Togo, several edible plants are used for food but also in the pharmacopoeia, notably X. mafaffa. Xanthosoma mafaffa is recognized as a food plant belonging to the Araceae family. This plant, originally native to Central and South America, is a recent (19th century) appearance in the tropical world. The name “New Cocoyam” reflects this late introduction into regions where the Xanthosoma genus, also known by various names such as Taro, Dasheen, and Eddoe, was already established14. The leaves as well as the tubers of this plant are commonly used in the diet of the Togolese population. Moreover, these leaves possess antimicrobial, antianemic, antiaflatoxin B1, and antiosteoporosis activities15,16. Furthermore, the chlorophylls a and b present in X. mafaffa leaves are powerful antioxidants that help to protect the body’s cells against cellular damage caused by free radicals17. These pigments are also known to detoxify the blood and cleanse the intestinal flora. The carotenoids, for them, are provitamins A. Dietary β-carotene and other provitamin A carotenoids such as α-carotene and cryptoxanthin can be obtained from several foods such as X. mafaffa18. The determination of biochemical compounds of X. mafaffa leaves was carried out using standard AOAC methods. The nutritional potential reported from X. mafaffa leaves is mainly attributed to the presence of essential amino acids such as threonine, valine, leucine, isoleucine, phenylalanine, and lysine19. The leaves of X. mafaffa are also rich in protein, lipids, carbohydrates, fibers, vitamins A and C, and minerals19. Among the minerals present in this plant, calcium, magnesium, iron, phosphorus, and zinc are present in appreciable quantities, along with traces of other minerals20. Previous studies reported that changes in various analysis conditions, such as the temperature, the harvesting time, the soil and age of the species, among other parameters, could significantly influence the content of these biochemical compounds21. Despite an extremely rich nutritional and phytochemical profile, taro leaves remain largely underutilized both due to a lack of awareness of their health benefits but also due to the presence of anti-nutritional factors, namely phytates and oxalates22.
The link between the chemical composition and antimicrobial activity of X. mafaffa leaf extract has not been explored to date. Hence, this study aims to identify the main biochemical compounds and to evaluate the antimicrobial activity of X. mafaffa leaves in order to make phytochemistry-bioactivity correlations and, finally, in order to better valorize this plant species.
MATERIALS AND METHODS
Vegetable material, reagents, equipment, and consumables: The X. mafaffa leaves (young and mature) were harvested in 2019 in Ecological Zone III at Ayome-Amou prefecture (Togo), with an age difference of 5 months at harvest time. This species was identified and conserved at the herbarium of the University of Lomé, Togo under number 15661. Once harvested, these leaves were then dried in the dark, under air conditioning at 20°C, for 14 days and ground using a knife mill (Brook Crompton, Ohio, USA) (Fig. 1).
The following reagents were purchased from Sigma-Aldrich (Steinheim, Germany): Catechin (C15H14O6) and sulfuric acid (H2SO4). All the other chemicals were purchased from Sigma-Aldrich (St. Louis, Missouri, USA): Tashiro indicator, ammoniacal iron (II) sulfate (NH4Fe (SO4)2), boric acid (H3BO3), and sodium hydroxide (NaOH).
Butanol, hydrochloric acid, methanol, hexane, and ethanol (Sigma-Aldrich) were of analytical grade. Water was purified by the Arum® 611 water purification system (Sartorius, Göttingen, Germany).
All spectrophotometry experiments were carried out using a spectrophotometer UV-visible (METASH, Waltham, USA).
Preparation of extracts: The powders of young leaves (YL) or mature leaves (ML) of X. mafaffa were macerated at a solid/liquid ratio (10% w/v) at ambient temperature for 72 hrs in hydroethanolic solvent of proportions in ethanol/water of 50/50 (v/v), further named 50%.
After maceration, the solutions were filtered through a filter paper (0.45 μm Whatman). Then, the filtrates obtained were dried by evaporation under vacuum using a Büchi rotary evaporator with the temperature of the water bath set at 45°C. The dry extracts were weighed and then stored in a small glass bottle, in a dark place, and the freezer for further analysis. At the end of this process, 2 extracts were collected for further analysis. The samples will be identified by the nature of the leaves, followed by the solvent used (i.e., YL-50% or ML-50%).

|
Phytochemical screening
Preliminary screening: The major chemical groups, such as alkaloids, flavonoids, tannins, saponins, coumarins, cardiac glucosides, terpenoids, coumarins, and total carbohydrates of each type of leaf, were determined by an approach based on solubility tests, staining, and/or precipitation reactions according to the methods described in the literature by Zhishen et al.23 and Malar and Chellaram24.
Determination of total proanthocyanidins: Proanthocyanidins are polyphenols derived from plants. The determination of proanthocyanidins was performed by the butanol/hydrochloric acid (BuOH/HCl) method described by Makkar et al.25. In a test tube, 0.2 mL of Ammoniacal iron sulfate solution (NH4Fe(SO4)2: 20 g/L) was introduced, followed by 7 mL of BuOH/HCl (95/5: v/v) and 0.2 mL of X. mafaffa extract solution (30 mg/mL). The mixture was incubated in a water bath at 95°C for 40 min to facilitate the formation of colored anthocyanidins. The pink or red coloration observed corresponding to the proanthocyanidin contents of the samples was read by UV-visible spectrophotometry, by optical density (OD) measurement at a wavelength λ = 550 nm, about catechin25.
The proanthocyanidin content (PC) was determined in each investigated sample and expressed as mg catechin equivalent (CE)/g of dry extract. The following formula was used to make the calculations (Eq. 1):
| (1) |
with OD = 0.280 equivalent to 1% w/v catechin.
Biochemical analysis
Water content analysis: The water content was determined using the method described by Vergun et al.26. The fresh X. mafaffa leaves in glass dishes were placed in a drying oven (STIK Instrument Equipment Ltd., Göttingen, Germany) and dried to reach a constant weight at 75 for 12 hrs. The weight of the leaves was measured before and after drying to determine their wet weight and the dry weight, respectively. The Sartorius BSA series precision balance (Sartorius, Göttingen, Germany) with a precision of 0.1 mg was used for weight measurement. During the measurement, a piece of white paper was placed under the dried leaves to prevent the broken leaves from affecting the next sample measurement results. The water content was determined according to the following formula (Eq. 2):
| (2) |
Where:
| Wfresh | = | Weight of fresh leaves expressed in grams | |
| Wdry | = | Weight of dry leaves expressed in grams |
Determination of total proteins: Proteins were determined by the Kjeldahl method described by Maehre et al.27 with slight modifications. The young or mature leaf powder (5 g) was dissolved in 10 mL of H2SO4 at 98%. After 6 hours of incubation at ambient temperature, the mineralization was distilled in the presence of 30% (w/v) sodium hydroxide. The distillate was collected in a beaker containing 10 mL of 4% (v/v) boric acid and 2 drops of Tashiro reagent and then titrated with 0.05 M sulfuric acid solution until the color turned pink. From the volume of sulfuric acid required determined previously, the total nitrogen content was determined according to the following formula (Eq. 3):
| (3) |
Where:
| N | = | Total nitrogen content expressed in percentage (percentage, g/100 g of dry matter) | |
| V | = | Volume of sulfuric acid solution (mL) required to obtain the color change | |
| C | = | Concentration of sulfuric acid solution used for titration (0.05 M) | |
| m | = | Sample mass (g) |
The nitrogen content obtained was converted into a percentage of crude protein by multiplying the result by a factor of 6.25.
Determination of total lipids: The total lipid content was determined using the method described by Vergun et al.26. The operation involved the extraction of lipids contained in 5 g of the investigated sample with hexane in a Soxhlet-type extractor (Gerhardt Analytical Systems, Königswinter, Germany). Then, the hexane was evaporated from the collected sample with a rotary evaporator (Model R-300, Büchi, Flawil, Switzerland), and the capsule was dried in an oven at 103°C to reach a constant weight. The weight difference before and after lipid extraction was used to obtain the total lipid content in g/100 g of sample, expressed by the following formula (Eq. 4):
| (4) |
Where:
| W0 | = | Weight of sample expressed in g (here 5 g) | |
| W1 | = | Weight of vial containing glass beads before extraction (without leave powder), expressed in grams | |
| W2 | = | Weight of the vial containing glass beads and fat after extraction from the leaf powder, expressed in grams |
Determination of total ashes: The experimental protocol used was described by Liu28 with some modifications. The total ash content was determined from 5 g of leaf powder. First, a porcelain crucible was heated at 550°C for 1 hr, then cooled in a desiccator and tared. Then, the leave powder (5 g) was added to the porcelain crucible, and the whole was progressively heated to 550°C in an oven for slow carbonization without ignition. The temperature was maintained at 550°C for 6 hrs to obtain white ash. The total ash content was calculated as follows (Eq. 5):
| (5) |
Where:
| W0 | = | Weight of empty incineration capsule expressed in grams | |
| W1 | = | Weight of incineration capsule loaded with sample before incineration, expressed in grams | |
| W2 | = | Weight of incineration capsule loaded with sample after incineration expressed in grams |
Determination of total fiber content: The total fiber content (TFC) was determined using the method described by Vergun et al.26. The young or mature leaf powder (1 g) was added to a volume of 200 mL of boiling H2SO4 at 0.25 N. After 30 min incubation at ambient temperature, the hydrolyzed mixture was filtered through the crucible, and the residue was rinsed with boiled distilled water to remove the acid from the filtrate recovered from inside the crucible. Again, 200 mL of boiled sodium hydroxide (NaOH, 0.313 N) was added to the crucible and boiled for an additional 30 min. The hydrolyzed samples were filtered again, and the residue was rinsed with boiled distilled water until the crucible was free of sodium hydroxide. The residue was rinsed again with a small amount of acetone and then drained. Finally, the residue in the crucible was dried in the oven at 105°C until reaching a constant weight. The crucible was placed in the muffle furnace at 550°C for 15 min and was burned completely. The crucible was then placed in the desiccator until a constant weight was achieved and calculated as (Eq. 6):
| (6) |
Where:
| W0 | = | Weight of empty incineration capsule expressed in grams | |
| W1 | = | Weight of incineration capsule loaded with sample before incineration, expressed in grams | |
| W2 | = | Weight of incineration capsule loaded with sample after incineration expressed in grams |
Determination of total digestible carbohydrates: The total digestible carbohydrate content (TDCC) was estimated by the difference method. It was calculated using the method of Menezes et al.29 according to the following formula (Eq. 7):
TDCC (%) = 100-(TPC+TLC+TAC) |
(7) |
Where:
| TDCC | = | Total digestible carbohydrate content expressed in percentage (g/100 g of dry matter) | |
| TPC | = | Total protein content expressed in percentage (g/100 g of dry matter) | |
| TLC | = | Total lipid content expressed in percentage (g/100 g of dry matter) | |
| TAC | = | Total crude ash content expressed in percentage (g/100 g of dry matter) |
The total digestible carbohydrate content (TDCC) plus the ash content (TAC) indicates the total carbohydrate content (TCC).
Determination of the energy value: The global energy value is the energy released by the combustion of macronutrients, i.e., proteins, carbohydrates, dietary fiber, and lipids contained in the diet, taking into account their coefficient of Atwater and Benedict, 4, 4, 1.91205, and 9 kcal, respectively30.
The global energy value expressed in Kilocalorie (kcal) was calculated from the following relationship (Eq. 8):
Energy value=100-(4×TPC+9×TLC+1.91205×TFC+4×TDCC) |
(8) |
Where:
| TPC | = | Total protein content expressed in percentage (g/100 g of dry matter) | |
| TLC | = | Total lipid content expressed in percentage (g/100 g of dry matter) | |
| TFC | = | Total fiber content expressed in percentage (g/100 g of dry matter) | |
| TDCC | = | Total digestible carbohydrate content expressed in percentage (g/100 g of dry matter) |
Determination of minerals: The mineral element analysis was carried out according to the AOAC methods with some modifications31,32. For sample preparation, the young or mature leaf powder (1 g) was digested by using 4 mL of H2SO4 at 98% and then 1 mL of H2O2 at 9% in a closed microwave system (Cem MARS Xpress, Matthews, USA). The volume of the digested samples was raised to 15 mL with deionized water and filtered through Whatman No. 42. The analysis of the filtered digested sample were carried out using Inductively Coupled Plasma-Atomic Emission Spectroscopy (ICP-AES) based on a quadrupole mass analyzer and octapole reaction system. The collision cell was used in He-mode for eliminating the possible polyatomic interferences. Likewise, the mineral elements P, K, Ca, and Fe were measured in He-mode, whereas the mineral elements Na and Mg were measured in a gas mode. The calibration solutions were prepared by the appropriate dilution of the single-element certified reference materials with 1.00±0.002 g/L for each element (Na, Mg, P, K, Ca, and Fe) with deionized water. Each plant sample was measured in triplicate by ICP-AES using Thermo Scientific™ Qtegra™ software for the analysis, with a peristaltic pump, a cross-flow nebulizer, and a ceramic central torch tube injector with an internal diameter of 2 mm.
Antimicrobial activity of X. mafaffa leaves
Microorganisms investigated: The antimicrobial activity of extracts of X. mafaffa was investigated against two Gram+ bacteria (i.e., Staphylococcus aureus ATCC 2921 and Streptococcus pneumoniae ATCC 49619), five Gram– bacteria strains (i.e., Pseudomonas aeruginosa ATCC 27853, Escherichia coli ATCC 25922, Shigella flexneiri ATCC 18593, Salmonella typhimirium ATCC 14028 and Klebsiella pneumoniae ATCC 700603) and also a yeast (i.e., Candida albicans ATCC 18331).
All strains were obtained from the Laboratory of Medical Bacteriology of the National Institute of Hygiene of Lomé, Togo.
Preparation of the inoculum: Bacterial and yeast cells were grown 18±2 hrs at 35°C in Mueller-Hinton broth (MHB). Then, they were centrifuged (3000 g, 5 min), and the collected pellets were diluted in 10 mL NaCl 0.90 % (w/v), allowing them to maintain the integrity of bacterial cells. The absorbance was measured by spectrophotometry and recorded at a wavelength of 625 nm with a plate reader (Thermo Multiskan FC). Bacterial and yeast cells were diluted in MHB to reach a 0.5 McFarland turbidity, which corresponds to 1 to 1.5×106 CFU/mL.
Determination of antimicrobial activity: The minimal inhibitory concentration (MIC), defined as the lowest concentration of extract with no visible bacterial growth, was determined using the microdilution tube method33.
The Mueller-Hinton broth was used to prepare one-half serial dilutions of concentrations ranging from 50 to 1 mg/mL. 10 μL of each bacterial or yeast inoculum was added to each tube containing 500 μL of the test solutions. Tubes without inoculum were considered as negative controls. All these tubes were incubated at 37°C for 24 hrs. The MIC was expressed here as the mg/mL of extract (w/v) in the tubes34.
For minimal bactericidal concentration (MBC) determination, 5 μL of each tube that did not show visible culture during MIC determination was removed and placed on nutrient agar for bacteria or yeast. After 24 hrs of incubation at 37°C, the lowest concentration of extract that did not give rise to colonies was taken as the MBC.
Finally, the MBC/MIC ratio was calculated to determine whether the antimicrobial effect observed was bactericidal or bacteriostatic. For an MBC/MIC ratio higher than 1, the antimicrobial effect of a substance is considered bacteriostatic. Inversely, when this ratio is less than or equal to 1, the antimicrobial effect is considered bactericidal. Finally, for an MBC/MIC ratio higher than or equal to 16, the antimicrobial activity is considered non-existent34.
Statistical analysis: All the experiments to quantify biochemical compounds and antimicrobial tests were carried out in triplicate. The results were expressed as the Mean±Standard Error of the Mean (SEM) of these independent experiments. Statistical significance was analyzed using Duncan’s Multiple Range Test (DMRT) in SPSS 11.5 Statistics Software (SPSS, Chicago, Illinois, USA) with a significance criterion set at p<0.05 compared to the respective control and between samples.
RESULTS AND DISCUSSION
Phytochemical screening
Qualitative screening findings: The qualitative phytochemical tests carried out on the different extracts allowed the detection of a variety of secondary metabolites, notably alkaloids, reducing compounds, flavonoids, tannins, cardiac glucosides, triterpenoids, saponins, total carbohydrates, and coumarins (Table 1).
Table 1 shows that young and mature leaves of X. mafaffa are rich in secondary metabolites, with the presence of eight families out of the nine investigated, which represents about 90% of the secondary metabolites searched. The young and mature leaves of X. mafaffa extracts contain similar phytochemical groups such as alkaloids, tannins, reducing compounds, total carbohydrates, saponins, triterpenoids, coumarins, and flavonoids. These molecules are concentrated in both young and mature leaves, due to the aerial part’s exposure to the same environmental stresses (e.g., UV, drought, temperature fluctuations). The use of a hydroethanolic solvent enables to obtain of extracts containing polar and semi-polar compounds such as tannins, triterpenoids, saponins, coumarins, and phenolic compounds. Yet, these compounds are well-known to be the main constituents reported to have antioxidant activities35. Current results are in agreement with those obtained by Dzotam et al.15, who conducted qualitative phytochemical screening on the 100% v/v methanolic extract of mature leaves of X. mafaffa. Former authors also evidenced the phenolic compounds, tannins, triterpenoids, saponins, coumarins, and sterols. Yet, contrary to current results, Dzotam et al.15 did not screen for the presence of alkaloids. Overall, the differences obtained could be due to the different solvents used (hydroethanolic solvent in current study vs methanol in Dzotam’s one), the harvesting period (June at the start of flowering in current study vs January at the end of flowering in that of Dzotam et al.15) and acclimatization (i.e., the region where the plant grew) seem to have influenced the chemical composition of this plant. By way of example, the differences observed between current work and that of Dzotam et al.15 could be linked to the choice of extraction solvent. Indeed, these authors’ studies focused on methanolic extracts, whereas current studies concerned hydroethanolic extracts. A large proportion of the secondary metabolites are defense molecules that ward off predators or inhibit the growth of competing plants.
The presence of these different chemical groups in the leaves of X. mafaffa is therefore a major asset, which justifies the therapeutic use of this food plant in Togo. Indeed, antibiotic activity could be due to plant alkaloids, while antioxidant, anti-inflammatory, and antiviral activities may derive from polyphenolic compounds35.
| Table 1: | Qualitative phytochemical screening of Xanthosoma mafaffa leaves | |||
| Analyzed metabolites | Identification reagents | YL-50% | ML-50% |
| Alkaloids | Dragendroff reagent | + | + |
| Wagner reagent | - | - | |
| Mayer reagent | + | + | |
| Tannins | Reaction with 1% w/v FeCI3 | + | + |
| Reaction with 10% w/v lead acetate | - | - | |
| Reducing compounds | Fehling’s test | + | + |
| Total carbohydrates | Molisch's test | + | + |
| Saponins | Froth test | + | + |
| Triterpenoids | Reaction with concentrated H2SO4 | + | + |
| Coumarins | Reaction with 10% w/v NaOH | + | + |
| Flavonoids | Reaction with 1% w/v NaOH | + | + |
| Cardiac glucosides | Reaction with CHCI3+H2SO4 | - | - |
| YL: Young leaves, ML: Mature leaves, 50%: 50/50 (v/v), Ethanol/water ratio (v/v), +: Presence of the analyzed compounds and -: Absence of the analyzed compounds | |||
|
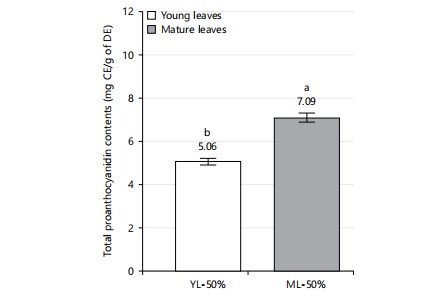
Total proanthocyanidin contents: To evaluate the effect of X. mafaffa leaves maturation on the concentration of proanthocyanidins, polyphenols derived from plants, a comparative analysis of extracts was carried out. Figure 2 shows the total proanthocyanidin content of the different extracts as a function of the maturity of the leaves (young or mature leaves).
The total proanthocyanidin content is determined at 5.1±0.2 and 7.1±0.2 mg CE/g dry extract for YL-50% and ML-50% extracts, respectively. Thus, the total proanthocyanidin content is significantly more concentrated (about 40% more) in mature leaves than in young leaves.
This can be attributed to the effect of species maturity on the chemical composition of the plant. Thus, the mature leaves of X. mafaffa are more interesting than the young leaves for further optimizing the extraction of the proanthocyanidins. Current results are in agreement with the study of Sharma et al.36, who reported that the concentration of proanthocyanidins depends strongly on the plant’s growth stage, but also the seasonal factors, the collection method in addition to the genetic and environmental factors. To the author’s knowledge, there are no articles reporting the investigation of proanthocyanins in X. mafaffa. Therefore, we have discussed current results due to their potential contribution to the understanding of proanthocyanins present in leafy vegetables. The total proanthocyanidin content determined in mature leaves (7.1±0.2 mg CE/g DE) is higher than that reported by Hernández-Jiménez et al.37 on the grape seeds of Vitis vinifera. Former authors reported a total proanthocyanidin content of 4.3 mg CE/g DE in the ethanolic extract, obtained by maceration at a solid/liquid ratio (10% w/v) for 24 hrs at 50°C of the powder of grape seeds of Vitis vinifera. Overall, the differences obtained could be linked to the choice of extraction process used (maceration with hydroethanolic solvent at a solid/liquid ratio of 10% w/v for 72 hrs at ambient temperature in current study vs simulated maceration in model wine solutions with varying ethanol concentrations for up to 10 days at room temperature in Hernández-Jiménez et al.37), the extraction time (72 hrs in current study vs 10 days in that of Hernández-Jiménez et al.37), and the nature of the species (X. mafaffa leaves in current study vs Vitis vinifera grape seeds in Hernández-Jiménez et al.37). In contrast, the total proanthocyanidin results are lower in current study than those reported by Ahammed et al.38 on methanolic extract of mature Grewia nervosa leaves from Bangladesh (45.8 CE/g DE). The differences in total proanthocyanidins obtained can be due to the different solvents used, the extraction process, the soil, climatic factors, and the chemical composition of the species36.
|
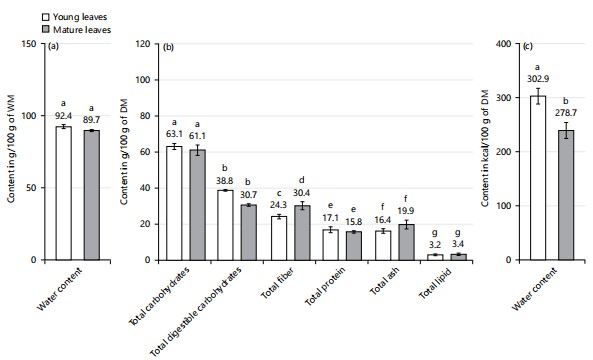
Biochemical analysis: Figure 3 shows the biochemical compounds in young and mature leaves of X. mafaffa. Upon maturation, the water content, the total carbohydrate, the digestible carbohydrate contents, and the total protein decrease between young leaves and mature leaves at the expense of the mature leaves. Yet, this decrease is only significant for the digestible carbohydrates. Inversely, the total fiber, the total ash, the total lipid, and the water content tend to increase with the leaves’ maturity, but not significantly.
Total water contents: The total water content (TWC), expressed in g/100 g of wet matter, is high in both leaves but slightly more important in young leaves compared to mature leaves (Fig. 3a). The water content determined is 92±1.4 and 89.7±0.7 g/100 g WM for young leaves and mature leaves, respectively. Yet, in current study, this difference in water content is not significant between YL-50 and ML-50%. According to the literature of de Almeida Jackix et al.39, the difference observed may be explained by the effect of maturity on the chemical composition of the species. Indeed, as the plant grows older, the accumulation of structural compounds such as cellulose and lignin, as well as the presence of secondary metabolites, is concomitant with the decrease in water content in X. mafaffa leaves.
Current results are in agreement with the study of Pant et al.40, who reported that the concentration of water depends strongly on the plant’s growth stage, but also the seasonal factors, the collection method in addition to the genetic and environmental factors. To the author's knowledge, no articles appear to report the determination of water content in fresh X. mafaffa leaves. Consequently, authors have discussed the results by comparing them with those obtained for other leafy vegetables. Besides, whatever the nature of the leaves, current results concerning total water contents present in X. mafaffa are superior to those obtained by Ahmed et al.41 on mature leaves of A. digitata (collected in Sudan; TWC: 8.9 g/100 g WM). Current results are also higher than those obtained by Ijarotimi et al.42 on mature M. oleifera leaves from Nigeria (TWC: 73 g/100 g WM). In addition to the plant sources, the differences in total water content obtained can be due to the soil, the climatic factors, the plant’s growth stage and the chemical composition of the species40. This result shows that X. mafaffa leaves can be classified as a water-rich leafy vegetable.
Total protein contents: Total protein content (TPC), expressed in g/100 g of dry matter, is slightly more concentrated in young leaves than in mature leaves, but the observed difference is not statistically significant (Fig. 3b). The total protein content determined is of 17.1±1.7 and 15.8±0.6 g/100 g DM for young leaves and mature leaves, respectively. This difference in the protein content observed in the different leaves can be attributed to the effect of the leaves maturity on the chemical composition of the species. Indeed, the growth of leaves requires active synthesis of cell membranes, enzymes, and other cellular components, which leads to a slight increase in the protein content of young leaves43. Moreover, the richness in protein content obtained for both types of X. mafaffa leaves suggests that they may be an interesting sources of proteins. The protein content of X. mafaffa leaves determined in current study is close to that obtained by Arogundade and Adedeji44 of mature X. mafaffa leaves (collected in Nigeria), reported a total protein content of 19.6 g/100 g DM. However, the TPC results determined for X. mafaffa in this study are about 2 times lower than those obtained by Ijarotimi et al.42 on mature M. oleifera leaves from Nigeria (TPC: 35.1 g/100 g DM). Overall, the differences obtained between current work and that of Ndong and colleagues could be linked to the nature of the species (mature leaves of X. mafaffa in current study vs mature leaves of Moringa oleifera in Ndong’s), the harvesting period (June at the start of flowering in current study vs December at the start of the dry season in Ndong’s) and acclimatization (i.e., the region where the plant was grown), which seem to have influenced the chemical composition of this plant. In general, proteins are essential macronutrients for various bodily functions, including tissue growth and repair, enzyme production, and immune function43. The leaves of X. mafaffa could serve as a sources of plant protein for local populations yet in addition to animal protein, in order to ensure amino acid balance.
Total lipid contents: Total lipid content (TLC), expressed in percentage of dry matter, seems more concentrated in mature leaves than in young leaves but yet, this difference is not significant Fig. 3b. The lipid content determined is 3.2±0.4 DM for young leaves, which is very close to 3.4±0.6 DM for mature leaves, respectively. The slightly higher bioavailability of lipids in mature leaves can be attributed to the effect of maturity on the chemical composition of the species. Indeed, the biosynthetic activity of lipids may be related notably to the activity of lipogenic enzymes and the incorporation of fatty acids into triglycerides during leaf maturity. Lipids are stored in vacuoles and cell membranes, contributing to energy reserve and cell structuring functions45. Furthermore, these two low lipid contents obtained suggest that the young and mature leaves of X. mafaffa can make a small contribution to lipid provision. In general, lipids are macromolecules essential for many biological functions, including cell membrane formation and energy storage45. Current results for lipid content in X. mafaffa leaves are 2 times lower than those obtained by Arogundade and Adedeji44 for mature X. mafaffa leaves (collected in Nigeria), with lipid content determined at 6.8 g/100 g DM. Likewise, current TLC results are 2 times lower than those obtained by Ijarotimi et al.42 on mature M. oleifera leaves from Nigeria (Total lipid content: 7.5 g/100 g DM). These low lipid contents suggest that X. mafaffa leaves can be consumed in antihypertensive diets but also by people with excess weight problems, with a contribution between 13.83 and 30.50% of the recommended dietary allowance (RDA) in lipids (Table 2) for a 70 kg adult.
| Table 2: | Contribution of organic and mineral substances of 100 g of dry matter of X. mafaffa leaves to the recommended daily allowance (RDA) | |||
| Elements analyzed | RDA for an adult (Male/Female) |
Contents in 100 g of d.w. of X. mafaffa leaves |
Contribution of 100 g of d.w. of X. mafaffa leaves to RDA (%) |
| Digestible carbohydrates (g) | 130a | 38.81 | 3.34 |
| Lipids (g) | 44-97a | 3.18 | 13.83-30.50 |
| Proteins (g) | 56 | 17.06 | 3.18 |
| Energy (kcal) | 2500/2000a | 302.9 | 8.25-6.60 |
| Na (mg) | 1500a | 329.8 | 4.54 |
| K (mg) | 4700b | 1927.01 | 2.43 |
| Ca (mg) | 900bc | 883.1 | 1.02 |
| Mg (mg) | 420a | 769.08 | 0.54 |
| P (mg) | 750bc | 396.3 | 1.98 |
| Fe (mg) | 27.40/58.80 | 11.02 | 2.49-5.33 |
| aReference nutrient intakes for minerals, carbohydrates, fiber, fat, proteins, and amino acids and b,cRecommended nutrient intakes for a body weight of 70 kg | |||
Total ash contents: Total ash content (TAC), expressed in g/100 g of dry matter, is slightly more concentrated in mature leaves than in young leaves but yet, but the difference is not statistically significant. The total ash content determined is 16.4±1.3 and 19.9±2.5 g/100 g DM for young leaves and mature leaves, respectively. Thus, the mature leaves of X. mafaffa are more beneficial than young leaves for mineral optimization. This bioaccumulation of minerals in mature leaves can be linked to the effect of maturity on the chemical composition of the species. Indeed, as the plant grows older, the minerals accumulate, in line with the membrane transport, leading to their storage in vacuoles, yet this phenomenon becomes more efficient during leaf maturity40. Besides, previous studies show that hormonal regulation and environmental stress signals can also play a dynamic role in mineral accumulation28. Note that the total ash content is linked to the presence of inorganic elements such as calcium, potassium, magnesium, iron, and other essential trace elements28. Current results are in agreement with the study of Pant et al.40, reporting that the concentration of ash depends strongly on the plant's growth stage, but also the seasonal factors, the collection method in addition to the genetic and environmental factors. The ash content results, determined here in X. mafaffa leaves, are also 3 times higher than those obtained by Ahmed et al.41, from mature leaves of A. digitata (collected in Sudan); a total ash content of 5.5 g/100 g DM. Likewise, the TAC results determined in the current study are also greater than those obtained by Ijarotimi et al.42 on mature M. oleifera leaves from Nigeria (TAC: 10.68 g/100 g DM). Thus, X. mafaffa leaves can be classified as an ash-rich leafy vegetable.
Total fiber content: The total fiber content (TFC), expressed in g/100 g of dry matter, is slightly more concentrated in mature leaves than in young leaves. The total fiber content determined is 24.3±1.3 and 30.4±2.2 g/100 g DM for young leaves and mature leaves, respectively. Yet, this difference between young leaves and mature leaves is not significant. This slight difference observed in the fiber content between young and mature leaves can be attributed to the effect of maturity on the chemical composition of the species. Indeed, some carbohydrates synthesized during the metabolic processes of photosynthesis can be converted into fibers as leaves grow older, which leads to an increase in fiber in mature leaves40. Previous studies by Liu28 reported that hormonal regulation and environmental stress signals can also play a complex role in fiber accumulation, influencing this dynamic positively or negatively, depending on the circumstances. Crude fibers are classified as carbohydrates that resist digestion by mammalian enzymes but can be broken down by rumen microorganisms. These fibers consist of cellulose, hemicellulose, lignin, and other soluble fibers28. Current results are in agreement with the study of Pant et al.40 reporting that the total fiber content depends strongly on the plant’s growth stage, but also the seasonal factors, the collection method in addition to the genetic and environmental factors.
Current results concerning the fiber contents of X. mafaffa leaves are also 3 times higher than those obtained by Pant et al.40 from mature leaves of A. digitata (collected in Ivory Coast); indeed, former authors reported at a total fiber content of 7.5 g/100 g DM41. Likewise, current TAC results are also greater than those reported by Ijarotimi et al.42 on mature M. oleifera leaves from Nigeria (4.68 g/100 g DM). Thus, the X. mafaffa leaves can be classified as an ash-rich leafy vegetable.
Total digestible carbohydrate: Kindly be informed that the carbohydrate content is inferred from the differing percentages identified in the proximate analysis, which accounts for total proteins, total fibers, total lipids, and ashes. Plant carbohydrates can be classified into two main types: Structural and non-structural carbohydrates. The structural carbohydrates are predominantly comprised of cellulose and hemicelluloses, forming the core of fibers in all plant tissues. Total digestible carbohydrate content (TDCC), expressed in g/100 g of dry matter, is slightly but significantly more concentrated in young leaves than in mature leaves. The total digestible carbohydrate content determined is 38.8±0.4 and 30.7±0.7 g/100 g DM for young leaves and mature leaves, respectively. Thus, the young leaves of X. mafaffa are more interesting than mature ones for the optimization of digestible carbohydrates. This high concentration of digestible carbohydrates in young leaves can be influenced by the effect of the maturity of the plant on the chemical composition of the species. Indeed, some of these carbohydrates can be converted into fibers or other forms of storage as the leaves grow older, which leads to their reduction in mature leaves39.
These highly digestible carbohydrate contents present in X. mafaffa suggest that both types of leaves can contribute considerably to carbohydrate intake, but with a higher contribution for young leaves. Current results for the digestible carbohydrate contents of X. mafaffa leaves are lower than those reported by Arogundade and Adedeji44 from mature X. mafaffa leaves collected in Nigeria, which reported a total digestible carbohydrate content of 51.3 g/100 g DM. However, current TDCC results are greater than those obtained by Ijarotimi et al.42 on mature M. oleifera leaves from Nigeria (TDCC: 35.7 g/100 g DM). In general, carbohydrates are a primary source of energy for the human body. Indeed, the digestible carbohydrates are broken down into monosaccharides such as glucose, which is then absorbed into the bloodstream and used by cells to produce ATP via glycolysis, the Krebs cycle, and the electron transport chain during digestion29. These compounds present in appreciable quantities in X. mafaffa leaves could be beneficial in diets requiring a rapid sources of energy45.
Total energy values: Figure 3c shows the metabolizable energy in young and mature leaves of X. mafaffa. The total energy value (TEV), expressed in kcal/100g of dry matter, is significantly higher in mature leaves compared to young leaves. Indeed, the total energy value determined is 302.9±14.3 and 278.7±14.9 kcal/100 g DM for young and mature leaves, respectively. This significant difference between YL and ML indicates a significant accumulation of energy in young mature leaves compared to mature leaves. The total energy value generally depends on the presence of macronutrients such as proteins, digestible carbohydrates, fibers, proteins, and other essential macronutrients. This bioaccumulation of certain macronutrients in young leaves can be attributed to the intense metabolic activity that takes place during their rapid growth phase. Note that leaf growth requires active synthesis of cell membranes, enzymes, and other cellular components, leading to an increase in the protein and digestible carbohydrate content of young leaves29. The total energy values obtained from the various X. mafaffa leaves are lower by about 15 to 22% than those obtained by Ijarotimi et al.42 from mature M. oleifera leaves collected in Nigeria. Indeed, former authors reported a total energy value of 358.7 kcal/100 g DM. This may be due to differences in the plant source. These greater energy value suggests that X. mafaffa leaves can be used as an energy source, with an important contribution to the recommended dietary allowance (RDA) in metabolizable energy (Table 2) for a 70 kg adult.
|
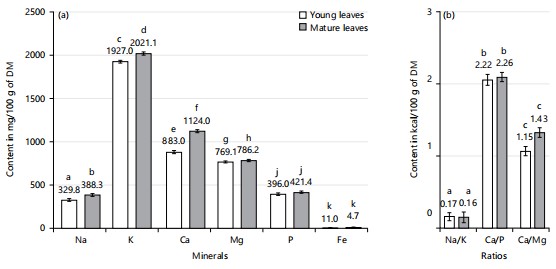
Total mineral contents: Figure 4a-b show the total mineral content (TMC) of young and mature X. mafaffa leaves. According to this figure, the Na, K, Ca, Mg, P, and Fe contents determined increase significantly with the leaves’ maturity. Indeed, the Total mineral content, expressed in mg/100 g DM, is significantly more concentrated in mature leaves than in young leaves. For young leaves, the contents of Na, K, Ca, Mg, P, and Fe are determined at 329.8±14.9, 1927.0±16.3, 883.1±17.5, 769.1±11.2, 396.3±14.1, and 11.0±1.8 g/100 g DM. For mature leaves, the contents of Na, K, Mg, Ca, P, and Fe obtained are higher, with values of 388.3±16.3, 2021.1±19.2, 1124.0±17.9, 786.2±12.3, 42.1±12.7, and 14.7±2.1 g/100 g DM, respectively. Thus, the maturity of X. mafaffa leaves favors a higher mineral concentration with a variation of 18, 5, 27, 2, 6, and 34% for Na, K, Ca, Mg, P, and Fe in favor of mature leaves.
This higher concentration of minerals in mature leaves compared to young leaves suggests that the plant mobilizes more minerals in mature leaves to support biochemical processes such as photosynthesis and resistance to environmental stresses46. Note that mineral levels are correlated with the plant’s specific needs, depending on its stage of growth. For example, potassium is essential for regulating stomatal opening and cell signal transmission, while calcium is involved in cell wall structuring47. However, the low iron content may also be due to the action of iron absorption inhibitors (e.g., polyphenols)42. Moreover, the mineral contents (i.e., Na, K, Mg, Ca, P, and Fe) determined in this study are lower than the values reported by Peluola-Adeyemi et al.20 for mature leaves of X. sagittifolium collected in Nigeria. Indeed, Peluola-Adeyemi et al.20 determined mineral contents of Na, K, Ca, Mg, P, and Fe of 19.3, 52.9, 17.7, 7.7, 44.4, and 39.3 mg/100 g DM, respectively.
The Na, K, Ca, Mg, P and Fe contents determined in current study are also higher than those obtained by Ijarotimi et al.42 on mature M. oleifera leaves from Nigeria with values of 70.9±0.5, 254.4±7.7, 423.2±25.9, 97.3±3.3, 301.7±45.1 and 21.7±0.6 mg/100 g dry matter, respectively. Note that the differences in total mineral content obtained can be due to the soil, the climatic factors, the plant’s growth stage, and the chemical composition of the species40. Consequently, the mineral content of both young and mature leaves of X. mafaffa is significant and can even be compared to that of certain vegetables and legumes recognized as major mineral sourcess, such as baobab, Moringa, and cassava leaves10. These minerals are vital elements necessary for the proper functioning and maintenance of the body.
| Table 3: | Minimum inhibitory concentrations, minimum bactericidal concentrations, and MBC/MIC ratio of extracts against the eight bacterial strains | |||
| Inhibition diameter (mm) | Sensitivity | ||||||
| Type | Germs | Positive control (ATB for bacteria and ATF for the yeast) |
YL-50% | ML-50% | MIC | MBC | MBC/ MIC |
| Gram– bacteria | E. coli | 14.60±0.10 | 0±0 | 0±0 | ND | ND | ND |
| S. flexneiri | 13.25±0.15 | 0±0 | 0±0 | ND | ND | ND | |
| S. typhimirium | 13.80±0.16 | 0±0 | 0±0 | ND | ND | ND | |
| K. pneumoniae | 14.22±0.20 | 12.25±0.35 | 0±0 | 12.5 | 50 | 4 | |
| P. aeruginosa | 14.15±0.15 | 0±0 | 0±0 | ND | ND | ND | |
| Gram+ ba`cteria | S. pneumoniae | 13.50±0.10 | 0±0 | 0±0 | ND | ND | ND |
| S. aureus | 13.10±0.15 | 0±0 | 0±0 | ND | ND | ND | |
| Yeast | C. albicans | 15.50±0.10 | 0±0 | 0±0 | ND | ND | ND |
| ND: Not Determined, ATB: Antibiotic (Gentamycin); ATF: Antifungal (Nystatin). YL: Young Leaves. ML: Mature Leaves. 50%: 50/50 (v/v) ethanol/water ratio. MIC: Minimum inhibitory concentrations (mg/mL), MBC: Minimum bactericidal concentrations (mg/mL) | |||||||
Mineral elements in humans are involved in a variety of functions, including those related to mineralization, the management of water balance, and the support of enzymatic systems. According to Razzaque and Wimalawansa48, they are also important constituents of the human diet by serving as enzyme cofactors for many physiological and metabolic processes. As a result, micronutrient deficiencies are serious health problems. Thus, iron (Fe), magnesium (Mg), and zinc (Zn) are fundamental minerals for human beings, especially for the production and oxygenation of blood cells, digestion, and overall blood circulation. Additionally, calcium (Ca) and potassium (K) are crucial for the process of muscle contraction9. In the case of sodium (Na), it is involved in the transmission of nerve impulses and the maintenance of fluid balance within the body47. Similarly, phosphorus (P) is crucial for energy storage mechanisms, notably in the form of ATP. These examples underscore the critical role of minerals in human health, emphasizing the need for sufficient intake to ensure proper physiological function. The presence of substantial mineral quantities in X. mafaffa leaves, therefore, constitutes a valuable nutritional asset. As such, their inclusion in the diet may offer benefits in terms of osteoporosis prevention, anti-aging effects, and immune system enhancement48. Moreover, the ratios of certain minerals were determined and are illustrated in Fig. 4b. Specifically, a Na/K ratio less than 1 is known to be beneficial for cardiovascular well-being49. The combination of low sodium levels and a significant presence of potassium further supports the use of these vegetables in diets aimed at managing hypertension48. In addition, the Ca/P and Ca/Mg ratios are greater than 1, consistent with the high mean calcium content found in the analyzed leaves. Therefore, these leafy vegetables demonstrate a richer calcium profile compared to phosphorus and magnesium. This elevated calcium content suggests that X. mafaffa leaves could be valuable additions to diets promoting skeletal health and pH balance through acid neutralization.
Antimicrobial activities of X. mafaffa leaves: The antimicrobial activity of X. mafaffa leaves was investigated on Gram-negative bacteria (S. aureus, S. flexneri, S. typhimurium, K. pneumoniae, and P. aeruginosa), Gram-positive bacteria (S. pneumoniae, and E. coli), and yeast (C. albicans). The minimum inhibitory concentrations (MIC), minimum bactericidal concentrations (MBC), and MBC/MIC ratios of X. mafaffa leaves extracts were determined against these eight bacterial strains and are reported in Table 3.
Among Gram-negative bacteria, the YL-50% extract has only an antimicrobial effect on K. pneumoniae, with an inhibition diameter of 12.25 mm.
In contrast, the extracts revealed no activity against Gram-positive bacteria or yeasts. Thus, the extracts YL-50% and ML-50% vs (S. aureus and S. pneumoniae) and extracts YL-50 and ML-50% vs C. albicans showed no antimicrobial activity.
On the whole of the results, the YL-50% extract has a more interesting antimicrobial activity than the ML-50% extract, yet, only against the K. pneumoniae. Thus, the YL-50% extract showed a MIC of 12.5 mg/mL and a BMC of 50 mg/mL on the K. pneumoniae strain. Moreover, the YL-50% extract exhibits a significant bacteriostatic activity against K. pneumoniae, as evidenced by the MBC/MIC ratio of 4. This selective antibacterial activity suggests a specific mechanism of action of YL-50% extract on the outer membrane of K. pneumoniae50.
At the exception of K. pneumoniae, current results are not in agreement with those of Gandhi et al.13, who found that the methanolic extract of X. mafaffa, obtained with a solid/liquid ratio of 10% (w/v), exhibited antimicrobial properties against Gram– bacteria such as Escherichia coli (MIC 512 μg/mL, MBC 1024 μg/mL, MBC/MIC ratio 2), Enterobacter aerogenes (MIC 512 μg/mL, MBC not data) and Providencia stuartii (MIC 512 μg/mL, MBC not data).
Correlations between the chemical composition and antimicrobial activity of the extract: The young and mature leaves of X. mafaffa contain a multitude of secondary metabolites. The extracts of young and mature X. mafaffa leaves contain similar phytochemical groups such as alkaloids, tannins, reducing compounds, total carbohydrates, saponins, triterpenoids, coumarins, and flavonoids (Table 1). Thus, the YL-50% extract exhibited appreciable K. pneumoniae antimicrobial activity, with a MIC determined at 12.5 mg/mL. This antimicrobial activity may be associated with the specific content of the extracted biomolecules. Indeed, although the two extracts contain similar classes of molecules, it is plausible that the extracted molecules differ not only in quantity but also in structure. These variations could thus explain the discrepancies observed in antimicrobial activity between these two extracts. Note that the chemical composition of X. mafaffa varies according to the species’ maturity51. In general, tannins are known for their ability to inhibit the growth of many microorganisms, including bacteria52. Flavonoids have antifungal and antibacterial properties52. Quinones and derivatives have antimicrobial properties38. Note that an MBC/MIC ratio is a key measure for assessing the efficacy of an antibiotic and guiding therapeutic choice. A low MBC/MIC ratio (≤4) indicates that the antibiotic is bactericidal, while a high ratio (>4) suggests that it is primarily bacteriostatic. For the YL-50% extract, the MBC/MIC ratio on the K. pneumoniae strain is 4. The MBC/MIC ratio obtained indicates that the hydroethanolic extract of young leaves has bacteriostatic activity on K. pneumoniae (MBC/MIC = 4). Consequently, young leaves can relieve illnesses such as pneumonia and urinary tract infections caused by K. pneumoniae.
CONCLUSION
This study focused on the leaves of X. mafaffa, commonly consumed as a leafy vegetable in many parts of the world, to evaluate their nutritional potential as well as their antimicrobial activity. The biochemical analysis carried out revealed that mature X. mafaffa leaves have a particularly remarkable nutritional composition, surpassing that of young leaves. They are rich in protein, fiber, easily digestible carbohydrates, and essential minerals such as calcium, magnesium, phosphorus, iron, and potassium. The microbiological tests carried out revealed significant antimicrobial activity of the YL-50% extract against K. pneumoniae. This activity is largely attributable to the bioactive compounds present in X. mafaffa leaves, such as flavonoids, tannins, and alkaloids, although their efficacy may vary according to leaf maturity. Given the nutritional quality of X. mafaffa leaves, these can be used as leafy vegetables capable of making a significant contribution to the dietary balance of populations. Their consumption could not only improve nutritional status but also offer protection against certain bacterial infections. This study provides solid evidence of X. mafaffa’s dual potential as a nutritious food sources and a natural antimicrobial agent. These results pave the way for further research aimed at optimizing the use of this plant, both in human nutrition and in the development of antimicrobial treatments. Their integration into public health strategies and the formulation of phytotherapeutic products represents a promising prospect for improving the health of populations.
SIGNIFICANCE STATEMENT
This study discovered the remarkable nutritional and antimicrobial potential of X. mafaffa leaves, which can be beneficial for improving dietary intake and combating bacterial infections. The findings highlight the rich composition of essential nutrients and bioactive compounds in mature leaves, demonstrating their potential role in both human nutrition and natural antimicrobial applications. This study will help researchers uncover the critical areas of bioactive compound efficacy and their influence on microbial resistance that many researchers were not able to explore. Thus, a new theory on the dual functionality of X. mafaffa as a nutritious food sources and a natural antimicrobial agent may be arrived at.
ACKNOWLEDGMENTS
The authors acknowledge the financial support provided by the Service of Cooperation and Cultural Action (SCCA) of Togo within the framework of the “Investing in the future” program implemented by the French Government. They also thank the Togolese Government's Direction of scholarships and internships for its financial support. Also, the authors thank the project of the “Lorraine Université d’Excellence” (Investissements d’avenir-ANR project number 15-004).
REFERENCES
- Baker, R.E., A.S. Mahmud, I.F. Miller, M. Rajeev and F. Rasambainarivo et al., 2022. Infectious disease in an era of global change. Nat. Rev. Microbiol., 20: 193-205.
- Hamdi, Y., I. Abdeljaoued-Tej, A.A. Zatchi, S. Abdelhak, S. Boubaker, J.S. Brown and A. Benkahla, 2021. Cancer in Africa: The untold story. Front. Oncol., 11.
- Iskandar, K., J. Murugaiyan, D.H. Halat, S. El Hage and V. Chibabhai et al., 2022. Antibiotic discovery and resistance: The chase and the race. Antibiotics, 11.
- Mancuso, G., A. Midiri, E. Gerace and C. Biondo, 2021. Bacterial antibiotic resistance: The most critical pathogens. Pathogens, 10.
- Akoua-Koffi, C., N. Guessennd, V. Gbonon, H. Faye-Ketté and M. Dosso, 2004. Methicillin-resistance of Staphylococcus aureus in Abidjan (1998-2001): A new hospital problem. Méd. Mal. Infectieuses, 34: 132-136.
- Dias, J.S., 2012. Nutritional quality and health benefits of vegetables: A review. Food Nutr. Sci., 3: 1354-1374.
- Nwobodo, D.C., M.C. Ugwu, C.O. Anie, M.T.S. Al‐Ouqaili, J.C. Ikem, U.V. Chigozie and M. Saki, 2022. Antibiotic resistance: The challenges and some emerging strategies for tackling a global menace. J. Clin. Lab. Anal., 36.
- Dangles, O., 2020. The antioxidant capacity of foods: Myths and reality. Cah. Nutr. Diet., 55: 176-183.
- Asad Ullah, S. Munir, S.L. Badshah, N. Khan and L. Ghani et al., 2020. Important flavonoids and their role as a therapeutic agent. Molecules, 25.
- Thiombiano, D.N.E., N. Lamien, D.S. Dibong, I.J. Boussim and B. Belem, 2012. The role of woody species in the management of the food shortage in Burkina Faso. Sci. Global Change Drought, 23: 86-93.
- Ferreira, P.M.P., D.F. Farias, J.T. de Abreu Oliveira and A. de Fátima Urano Carvalho, 2008. Moringa oleifera: Bioactive compounds and nutritional potential. Rev. Nutr., 21: 431-437.
- Pareek, A., M. Pant, M.M. Gupta, P. Kashania and Y. Ratan et al., 2023. Moringa oleifera: An updated comprehensive review of its pharmacological activities, ethnomedicinal, phytopharmaceutical formulation, clinical, phytochemical, and toxicological aspects. Int. J. Mol. Sci., 24.
- Gandhi, G.R., S. Ignacimuthu, M.G. Paulraj and P. Sasikumar, 2011. Antihyperglycemic activity and antidiabetic effect of methyl caffeate isolated from Solanum torvum Swartz. fruit in streptozotocin induced diabetic rats. Eur. J. Pharmacol., 670: 623-631.
- Onwueme, I.C., 1978. The Tropical Tuber Crops: Yams, Cassava, Sweet Potato and Cocoyams. John Wiley and Sons, New York, USA, ISBN:9780471996088, Pages: 234.
- Dzotam, J.K., F.K. Touani and V. Kuete, 2016. Antibacterial and antibiotic-modifying activities of three food plants (Xanthosoma mafaffa Lam., Moringa oleifera (L.) schott and Passiflora edulis Sims) against multidrug-resistant (MDR) Gram-negative bacteria. BMC Complementary Altern. Med., 16.
- Boakye, A.A., F.D. Wireko-Manu, I. Oduro, W.O. Ellis, M. Gudjónsdóttir and I.S. Chronakis, 2018. Utilizing cocoyam (Xanthosoma sagittifolium) for food and nutrition security: A review. Food Sci. Nutr., 6: 703-713.
- Martins, T., A.N. Barros, E. Rosa and L. Antunes, 2023. Enhancing health benefits through chlorophylls and chlorophyll-rich agro-food: A comprehensive review. Molecules, 28.
- Novidzro, K.M., K. Wokpor, B.A. Fagla, K. Koudouvo, K. Dotse, E. Osseyi and K.H. Koumaglo, 2019. Study of some physicochemical parameters and analysis of mineral elements, chlorophyllian pigments and carotenoids of Griffonia simplicifolia seed oil [In French]. Int. J. Biol. Chem. Sci., 13: 2360-2373.
- Wallace, P.A., E.K. Marfo and W.A. Plahar, 1998. Nutritional quality and antinutritional composition of four non-conventional leafy vegetables. Food Chem., 61: 287-291.
- Peluola-Adeyemi, O.A., M.A. Idowu, L.O. Sanni and G.J. Bodunde, 2014. Effect of some extrusion parameters on the nutrient composition and quality of a snack developed from cocoyam (Xanthosoma sagittifolium) flour. Afr. J. Food Sci., 8: 510-518.
- Rosinger, C. and M. Bonkowski, 2021. Soil age and soil organic carbon content shape biochemical responses to multiple freeze-thaw events in soils along a postmining agricultural chronosequence. Biogeochemistry, 155: 113-125.
- Mitharwal, S., A. Kumar, K. Chauhan and N.K. Taneja, 2022. Nutritional, phytochemical composition and potential health benefits of taro (Colocasia esculenta L.) leaves: A review. Food Chem., 383.
- Zhishen, J., T. Mengcheng and W. Jianming, 1999. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem., 64: 555-559.
- Malar, C.G.R. and C. Chellaram, 2018. Phytochemical screening, quantification of total phenols, total flavonoids and antimicrobial activity of stem extracts of Salacia oblonga. Indian J. Sci. Technol., 11.
- Makkar, H.P.S., G. Gamble and K. Becker, 1999. Limitation of the butanol-hydrochloric acid-iron assay for bound condensed tannins. Food Chem., 66: 129-133.
- Vergun, O., D. Rakhmetov, S. Rakhmetova and V. Fishchenko, 2022. Comparative study of biochemical composition of Paulownia tomentosa (Thunb.) Steud. genotypes. Agrobiodiversity Improving Nutr. Health Life Qual., 6: 180-190.
- Mæhre, H.K., L. Dalheim, G.K. Edvinsen, E.O. Elvevoll and I.J. Jensen, 2018. Protein determination-method matters. Foods, 7.
- Liu, K., 2019. Effects of sample size, dry ashing temperature and duration on determination of ash content in algae and other biomass. Algal Res., 40.
- Menezes, E.W., A.T. de Melo, G.H. Lima and F.M. Lajolo, 2004. Measurement of carbohydrate components and their impact on energy value of foods. J. Food Compos. Anal., 17: 331-338.
- Atwater, W.O. and F.G. Benedict, 1902. Experiments on the Metabolism of Matter and Energy in the Human Body, 1898-1900. U.S. Government Printing Office, Washington, DC, United State, Pages: 147.
- Ndomou, S.C.H., F.T. Djikeng, G.B. Teboukeu, H.T. Doungue, H.A.K. Foffe, C.T. Tiwo and H.M. Womeni, 2021. Nutritional value, phytochemical content, and antioxidant activity of three phytobiotic plants from West Cameroon. J. Agric. Food Res., 3.
- Jarošová, M., D. Milde and M. Kuba, 2014. Elemental analysis of coffee: A comparison of ICP-MS and AAS methods. Czech J. Food Sci., 32: 354-359.
- Chebaibi, A., Z. Marouf, F. Rhazi-Filali, M. Fahim and A. Ed-Dra, 2016. Evaluation of antimicrobial activity of essential oils from seven Moroccan medicinal plants. Phytothérapie, 14: 355-362.
- Olajuyigbe, A.A., O.O. Olajuyigbe and R.M. Coopoosamy, 2020. Interaction of Ziziphus mucronata subsp. mucronata methanol extract and first-line antibiotics is synergistic in vitro through production of reactive oxygen species. J. Trop. Med., 2020.
- Kadum, H., A. Abdul Hamid, F. Abas, N.S. Ramli, A.K.S. Mohammed, B.J. Muhialdin and A.H. Jaafar, 2019. Bioactive compounds responsible for antioxidant activity of different varieties of date (Phoenix dactylifera L.) elucidated by 1H-NMR based metabolomics. Int. J. Food Prop., 22: 462-476.
- Sharma, A., B. Shahzad, Abdul Rehman, R. Bhardwaj, M. Landi and B. Zheng, 2019. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules, 24.
- Hernández-Jiménez, A., J.A. Kennedy, A.B. Bautista-Ortín and E. Gómez-Plaza, 2012. Effect of ethanol on grape seed proanthocyanidin extraction. Am. J. Enol. Vitic., 63: 57-61.
- Ahammed, M.S., M. Shariful Islam, M. Mosiqur Rahman, S.F. Koly and K. Rakib et al., 2018. In vitro antioxidant and cholinesterase inhibitory activities of methanolic extract of Grewia nervosa L. (Family: Tiliaceae) leaf. Eur. J. Med. Plants, 25.
- de Almeida Jackix, E., E.B. Monteiro, H.F. Raposo, E.C. Vanzela and J. Amaya-Farfán, 2013. Taioba (Xanthosoma sagittifolium) leaves: Nutrient composition and physiological effects on healthy rats. J. Food Sci., 78: H1929-H1934.
- Pant, P., S. Pandey and S. Dall'Acqua, 2021. The influence of environmental conditions on secondary metabolites in medicinal plants: A literature review. Chem. Biodivers., 18.
- Ahmed, A.E.M., M. Mozzon, A.B.A. Dawod, E.O.A. Mustafa and S.A. Mukarram et al., 2024. Evaluation of the nutritional impact of baobab leaves (Adansonia digitata L.) as a dietary intervention to combat nutrient deficiencies and poverty-related health problems. Nutrients, 16.
- Ijarotimi, O.S., O.A. Adeoti and O. Ariyo, 2013. Comparative study on nutrient composition, phytochemical, and functional characteristics of raw, germinated, and fermented Moringa oleifera seed flour. Food Sci. Nutr., 1: 452-463.
- Yu, Y., M. Kleuter, S.T. Dinani, L.M. Trindade and A.J. van der Goot, 2023. The role of plant age and leaf position on protein extraction and phenolic compounds removal from tomato (Solanum lycopersicum) leaves using food-grade solvents. Food Chem., 406.
- Arogundade, O.O. and O. Adedeji, 2020. The importance of nutritive parameters in the taxonomy of some corm-producing members of the family Araceae. Notulae Sci. Biol., 12: 318-333.
- van der Meer-Janssen, Y.P.M., J. van Galen, J.J. Batenburg and J.B. Helms, 2010. Lipids in host-pathogen interactions: Pathogens exploit the complexity of the host cell lipidome. Prog. Lipid Res., 49: 1-26.
- Ahmed, N., B. Zhang, Z. Chachar, J. Li and G. Xiao et al., 2024. Micronutrients and their effects on horticultural crop quality, productivity and sustainability. Sci. Hortic., 323.
- Zhang, L., D. Li, Y. Yao and S. Zhang, 2020. H2O2, Ca2+, and K+ in subsidiary cells of maize leaves are involved in regulatory signaling of stomatal movement. Plant Physiol. Biochem., 152: 243-251.
- Razzaque, M.S. and S.J. Wimalawansa, 2025. Minerals and human health: From deficiency to toxicity. Nutrients, 17.
- Al-Jameil, N., F.A. Khan, M.F. Khan and H. Tabassum, 2014. A brief overview of preeclampsia. J. Clin. Med. Res., 6: 1-7.
- Anek, P., S. Kumpangcum, S. Roytrakul, C. Khanongnuch, C. Saenjum and K. Phannachet, 2024. Antibacterial activities of phenolic compounds in Miang extract: Growth inhibition and change in protein expression of extensively drug-resistant Klebsiella pneumoniae. Antibiotics, 13.
- Ngom, S., M. Diop, M. Mbengue, F. Faye, J.M. Kornprobst and A. Samb, 2014. Chemical composition and antimicrobial properties of the essential oils of Ocimumbasilicum and Hyptis suaveolens harvested from Dakar Region in Senegal [In French]. Afrique Sci.: Revue Int. des Sci. Technologie, 10: 109-117.
- Manach, C., A. Scalbert, C. Morand, C. Remesy and L. Jimenez, 2004. Polyphenols: Food sources and bioavailability. J. Clin. Nutr., 79: 727-747.
How to Cite this paper?
APA-7 Style
Prospère,
K.E., Mawuéna,
N.K., Kokouvi,
D., Honoré,
K.K., Laetitia,
C. (2025). Phytochemicals, Nutritional Value, and Antimicrobial Properties of Xanthosoma mafaffa. Research Journal of Phytochemistry, 19(1), 38-57. https://doi.org/10.3923/rjp.2025.38.57
ACS Style
Prospère,
K.E.; Mawuéna,
N.K.; Kokouvi,
D.; Honoré,
K.K.; Laetitia,
C. Phytochemicals, Nutritional Value, and Antimicrobial Properties of Xanthosoma mafaffa. Res. J. Phytochem 2025, 19, 38-57. https://doi.org/10.3923/rjp.2025.38.57
AMA Style
Prospère
KE, Mawuéna
NK, Kokouvi
D, Honoré
KK, Laetitia
C. Phytochemicals, Nutritional Value, and Antimicrobial Properties of Xanthosoma mafaffa. Research Journal of Phytochemistry. 2025; 19(1): 38-57. https://doi.org/10.3923/rjp.2025.38.57
Chicago/Turabian Style
Prospère, Kanabiya, Essodjolon, Novidzro Kosi Mawuéna, Dotse Kokouvi, Koumaglo Kossi Honoré, and Canabady-Rochelle Laetitia.
2025. "Phytochemicals, Nutritional Value, and Antimicrobial Properties of Xanthosoma mafaffa" Research Journal of Phytochemistry 19, no. 1: 38-57. https://doi.org/10.3923/rjp.2025.38.57

This work is licensed under a Creative Commons Attribution 4.0 International License.